ОСНОВНЫЕ СВОЙСТВА НЕЙРОНОВ
Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
Механизм
проведения возбуждения по нервным
волокнам зависит от их типа. Существуют
два типа нервных волокон: миелиновые и
безмиелиновые.
Процессы
метаболизма в безмиелиновых волокнах
не обеспечивают быструю компенсацию
расхода энергии. Распространение
возбуждения будет идти с постепенным
затуханием – с декрементом. Декрементное
поведение возбуждения характерно для
низкоорганизованной нервной системы.
Возбуждение распространяется за счет
малых круговых токов, которые возникают
внутрь волокна или в окружающую его
жидкость. Между возбужденными и
невозбужденными участками возникает
разность потенциалов, которая способствует
возникновению круговых токов. Ток будет
распространяться от «+» заряда к «—».
В месте выхода кругового тока повышается
проницаемость плазматической мембраны
для ионов Na, в результате чего происходит
деполяризация мембраны. Между вновь
возбужденным участком и соседним
невозбужденным вновь возникает разность
потенциалов, что приводит к возникновению
круговых токов. Возбуждение постепенно
охватывает соседние участки осевого
цилиндра и так распространяется до
конца аксона.
В
миелиновых волокнах благодаря совершенству
метаболизма возбуждение проходит, не
затухая, без декремента. За счет большого
радиуса нервного волокна, обусловленного
миелиновой оболочкой, электрический
ток может входить и выходить из волокна
только в области перехвата. При нанесения
раздражения возникает деполяризация
в области перехвата А, соседний перехват
В в это время поляризован. Между
перехватами возникает разность
потенциалов, и появляются круговые
токи. За счет круговых токов возбуждаются
другие перехваты, при этом возбуждение
распространяется сальтаторно,
скачкообразно от одного перехвата к
другому. Сальтаторный способ распространения
возбуждения экономичен, и скорость
распространения возбуждения гораздо
выше (70—120 м/с), чем по безмиелиновым
нервным волокнам (0,5–2 м/с).
Существует
три закона проведения раздражения по
нервному волокну.
Закон
анатомо-физиологической целостности.
Проведение
импульсов по нервному волокну возможно
лишь в том случае, если не нарушена его
целостность. При нарушении физиологических
свойств нервного волокна путем охлаждения,
применения различных наркотических
средств, сдавливания, а также порезами
и повреждениями анатомической целостности
проведение нервного импульса по нему
будет невозможно.
Закон
изолированного проведения возбуждения.
Существует
ряд особенностей распространения
возбуждения в периферических, мякотных
и безмякотных нервных волокнах.
В
периферических нервных волокнах
возбуждение передается только вдоль
нервного волокна, но не передается на
соседние, которые находятся в одном и
том же нервном стволе.
В
мякотных нервных волокнах роль изолятора
выполняет миелиновая оболочка. За счет
миелина увеличивается удельное
сопротивление и происходит уменьшение
электрической емкости оболочки.
В
безмякотных нервных волокнах возбуждение
передается изолированно. Это объясняется
тем, что сопротивление жидкости, которая
заполняет межклеточные щели, значительно
ниже сопротивления мембраны нервных
волокон. Поэтому ток, возникающий между
деполяризованным участком и
неполяризованным, проходит по межклеточным
щелям и не заходит при этом в соседние
нервные волокна.
Закон
двустороннего проведения возбуждения.
Нервное
волокно проводит нервные импульсы в
двух направлениях – центростремительно
и центробежно.
В
живом организме возбуждение проводится
только в одном направлении. Двусторонняя
проводимость нервного волокна ограничена
в организме местом возникновения
импульса и клапанным свойством синапсов,
которое заключается в возможности
проведения возбуждения только в одном
направлении.
Развитие и рост нейронов
Современные ученые до сих пор дискутируют на тему деления нервных клеток, т.к. единого мнения по этому вопросу в сфере анатомии на данный момент нет. Многие специалисты в этой области уделяют больше внимания свойствам, а не строению нейронов, что является более важным и актуальным вопросом для современной науки.
Наиболее распространенная версия – развитие нейрона происходит из клетки, деление которой прекращается еще до момента выпуска отростков. Сначала развивается аксон, после чего дендриты.
Зависимо от основного функционала, места расположения и степени активности, нервные клетки развиваются по-разному. Их размеры существенно варьируются в зависимости от места расположения и выполняемых функций.
По характеру пропускаемых ионов:
а) каливые
б)натривые
в)кальцивые
г)хлорные
3. По скорости
инактивации,
т.е. закрывания:
а) быстроинактивируюшиеся,
т.е. быстро переходящие в закрытое
состояние. Они обеспечивают быстро
нарастающее снижение МП и такое же
быстрое восстановление.
б) медленноинактирующиеся.
Их открывание вызывает медленное
снижение МП и медленное его восстановление.
4. По механизмам
открывания:
а) потенциалзависямые,
т.е. те, которые открываются при
определенном уровне потенциала мембраны.
б) хемозависимые,
открывающиеся при воздействии на
хеморецепторы мембраны клетки
физиологически активных веществ
(нейромедиаторов. гормонов и т. д).
В настоящее время
установлено, что ионные каналы имеют
следующее строение:
1
.Селективный фильтр, расположенный в
устье канала. Он обеспечивает прохождение
через канал строго
определенных
ионов.
2.Активационные
ворота, которые открываются при
определенном уровне мембранного
потенциала или действии соответствующего
ФАВ. Активационные ворота потенциалзависямых
каналов имеется сенсор, который открывает
их на определенном уровне МП.
3.Инактивационные
ворота, обеспечивающие закрывания
канала и прекращение проведения ионов
по каналу на определенном уровне МП.
Неспецифические ионные каналы не имеют
ворот.
Активный
транспорт
осуществляется с помощью энергии АТФ.
К этой группе транспортных систем
относятся натрий-калнезый насос,
кальциевый насос, хлорный насос.
Пассивный
транспорт.
Передвижение ионов осуществляется по
градиенту концентрации без затрат
энергии. Например, вход калия в клетку
и выход из неё по калиевым каналам.
Сопряженный
транспорт.
Противоградиентный перенос ионов без
затрат энергии. Например таким образом
происходит натрий натриевый,
натрий-кальциевый, калий -калиевый обмен
ионов. Он происходит за счет разности
концентрации других ионов.
Механизм проведения возбуждения по нерву.
Возбуждение
(потенциал действия — ПД) распространяется
в аксонах, телах нервных клеток, а также
иногда в дендритах без снижения амплитуды
и без снижения скорости (бездекрементно).
Механизм распространения возбуждения
у различных нервных волокон неодинаков.
При распространении возбуждения по
безмиелиновому нервному волокну механизм
проведения включает два компонента:
раздражающее действие катэлектротона,
порождаемое локальным ПД, на соседний
участок электровозбудимой мембраны и
возникновение ПД в этом участке мембраны.
Локальная деполяризация мембраны
нарушает электрическую стабильность
мембраны, различная величина поляризации
мембраны в смежных ее участках порождает
электродвижущую силу и местный
электрический ток, силовые линии
которого замыкаются через ионные каналы.
Активация ионного канала повышает
натриевую проводимость, после
электротонического достижения
критического уровня деполяризации
(КУД) в новом участке мембраны генерируется
ПД. В свою очередь этот потенциал действия
вызывает местные токи, а они в новом
участке мембраны генерируют потенциал
действия. На всем протяжении нервного
волокна происходит процесс новой
генерации потенциала действия мембраны
волокна. Данный тип передачи возбуждения
называется непрерывным.
Скорость
распространения возбуждения пропорциональна
толщине волокна и обратно пропорциональна
сопротивлению среды. Проведение
возбуждения зависит от соотношения
амплитуды ПД и величины порогового
потенциала. Этот показатель называется
гарантийный фактор(ГФ) и равен 5 —
7, т.е. ПД должен быть выше порогового
потенциала в 5- 7 раз. Если ГФ = 1 проведение
ненадёжно, если ГФ
Наличие у
миелиновых волоконоболочки, обладающей
высоким электрическим сопротивлением,
а также участков волокна, лишенных
оболочки — перехватов Ранвье создают
условия для качественно нового типа
проведения возбуждения по миелиновым
нервным волокнам. Вмиелинизированном
волокне токи проводятся только в
зонах, не покрытых миелином, — перехватах
Ранвье, в этих участках и генерируется
очередной ПД. Перехваты длиной 1 мкм
расположены через 1000 — 2000 мкм, характеризуются
высокой плотностью ионных каналов,
высокой электропроводностью и низким
сопротивлением. Распространение ПД в
миэлинизированных нервных волокнах
осуществляетсясальтаторно—
скачкообразно от перехвата к перехвату,
т.е. возбуждение (ПД) как бы «перепрыгивает»
через участки нервного волокна, покрытые
миелином, от одного перехвата к другому.
Скорость такого способа проведения
возбуждения значительно выше, и он более
экономичен по сравнению с непрерывным
проведением возбуждения, поскольку в
состояние активности вовлекается не
вся мембрана, а только ее небольшие
участки в области перехватов, благодаря
чему уменьшается нагрузка на ионный
насос.

Схема распространения
возбуждения в безмиелиновых и миелиновых
нервных волокнах.
8 Потенциал действия, его фазы, их происхождение.
Потенциал
действия
— это быстрое колебание мембранного
потенциала возникающее при возбуждении
мембраны.
Фазы:
1)
медленная деполяризация
(так же локальный ответ) — возникает
вследствие увеличение проницаемости
мембраны для ионов натрия. Под пороговый
стимул недостаточен, чтобы вызвать
быструю деполяризацию сразу. Длительность
фазы зависит от силы раздражителя.
2)
быстрая деполяризация
— характеризуется быстрым уменьшением
мембранного потенциала и даже перезарядкой
мембраны (овершут): внутренняя ее часть
на некоторое время становится заряженной
положительно, а внешняя отрицательно.
Это происходит вследствие лавинообразно
по ступающего натрия внутрь клетки. В
отличие от локального ответа скорость
и величина деполяризации не зависит от
силы раздражителя. Продолжительность
фазы деполяризации в нервном волокне
лягушки составляет около 0.2 — 0.5 мс.
3)
реполяризация
(продолжительность 0.5-0.8 мс) — мембранный
потенциал постепенно восстанавливается
и достигает 75 — 85% потенциала покоя.
2
и 3 фазы называются пиком потенциала
действия.
4)
следовая деполяризация —
является продолжением фазы реполяризации
и характеризуется более медленным (по
сравнению с фазой реполяризации)
восстановлением потенциала покоя
5)
следовая гиперполяризация
— представляет собой временное увеличение
мембранного потенциала выше исходного
уровня.
4
и 5 фазу называют следовыми явлениями
Причины и механизмы расстройств возбудимости и проводимости нервной клетки
Гипоксемия,
отравление цианидами, динитрофенолом,
азидами и другими токсическими веществами
приводит к нарушению метаболизма и
уменьшению выработки энергии в нервной
клетке, что резко угнетает возбудимость
нейронов. Дефицит тиамина, цианокобаламина,
сдавление или охлаждение нерва,
воспаление, гипоксия, действие некоторых
ядов и токсинов микроорганизмов нарушает
способность проводить нервное возбуждение
и часто приводит к нейродистрофическим
процессам, в частности, в миелиновой
оболочке нерва. Ведущими механизмами
нарушения возбудимости и проводимости
являются изменения мембранного
потенциала, генерации и проведения
потенциала действия по мембранам
нейрона. Они проявляются изменениями
деятельности селективных каналов
мембран, нарушениями их проницаемости
для различных ионов, а также возбудимости
клеточной мембраны.
Изменения
свойств нейронов при действии повреждающего
фактора развиваются фазно.
После
воздействия раздражителя нервная клетка
сначала, не теряя структурной целостности,
прекращает свое функционирование, а
при усилении повреждающего
действия нейрон погибает. Если же этот
агент устраняется вовремя, то нервная
ткань способна возвращаться в нормальное
состояние.
Обратимое
фазное изменение возбудимости и
проводимости возбудимой ткани, возникающее
под влиянием сильного раздражения
получило название парабиоза.
В зависимости от степени повреждения
нервных волокон различают несколько
фаз парабиоза. При небольшом повреждении
нерва в ответ на сильное или слабое
раздражение иннервируемая им мышца
отвечает одинаковыми по силе сокращениями.
Этоуравнительная
фаза.
По
мере углубления альтерации нерва
возникает парадоксальная
фаза,
т.е. в ответ на сильное раздражение нерва
мышца отвечает слабыми сокращениями,
в то время как умеренные по силе
раздражения вызывают более энергичные
мышечные сокращения. В последней фазе
парабиоза — фазе
торможения
—
никакие раздражения нерва не способны
вызвать мышечное сокращение. Учение о
парабиозе, разработанное Н.Е. Введенским
(1901), широко используется в неврологии.
Например, при повреждении нерва с полной
утратой его связи с телом нейрона
происходит дегенерация нервных волокон.
Через несколько дней после начала
дегенерации нерв утрачивает возбудимость.
При частичном повреждении периферического
нерва изменения возбудимости в направлении
нейрона минимальные, а в дальнейшем
происходит регенерация нервных волокон.
При повреждении
в ЦНС нервное волокно дегенерирует в
направлении нейрона, который чаще всего
погибает. Расстройства возбудимости
(инертность, запредельное торможение)
могут быть причиной возникновения
неврологических и психических расстройств
(маниакальных и судорожных состояний,
галлюцинаций, бреда).
Особенности строения
Структура включает в себя три основных составляющих:
- Тело. Тело включает в себя нейроплазму, ядро, которое разграничено мембранным веществом. Хромосомы ядра содержат гены, отвечающие за кодировку синтеза белков. Здесь также осуществляется синтез пептидов, которые требуются для обеспечения нормальной работы отростков. Если тело будет повреждено, то в скором времени произойдет и разрушение отростков. При повреждении одного из отростков (при условии сохранения целостности тела) он будет постепенно регенерироваться.
- Дендриты. Образуют дендритное дерево, имеют безграничное число синапсов, сформированных аксонами и дендритами соседних клеток.
- Аксон. Отросток, который, кроме нейронов, не встречается больше ни в одних клетках. Сложно переоценить их значение (например, аксоны ганглиозных клеток ответственны за формирование зрительного нерва).
Классификация нейронов в соответствии с функциональными и морфологическими признаками выглядит следующим образом:
- по числу отростков.
- по типу взаимодействия с другими клетками.
Все нейроны получают грандиозное число электрических импульсов из-за наличия множества синапсов, которые расположены по всей поверхности нейронной структуры. Импульсы также получаются через молекулярные рецепторы ядра. Электрические импульсы передаются разными нейромедиаторами и модуляторами. Поэтому важным функционалом также можно считать способность интеграции полученных сигналов.
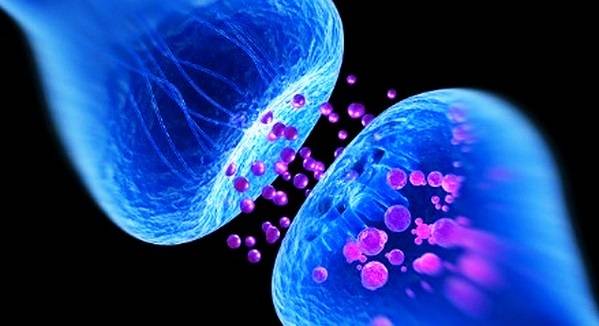
Чаще всего сигналы интегрируются и обрабатываются в синапсах, после чего в остальных частях нейронной структуры суммируются постсинаптические потенциалы.